Нейрофизиологический и формально-логический базис нейроподобных вычислений
4.2. Базовые положения теории функциональных систем
(Нейро)физиология по своему научному статусу интегрирует все достижения фундаментальных исследований физических, биофизических, химических, биохимических, молекулярно-биологических и нейрофизиологических процессов, протекающих в реальных нейронах, корковых и подкорковых образованиях, в периферической, вегетативной и центральной нервной системе и других нервных образованиях, которые участвуют в формировании устойчивых целенаправленных форм поведения и приспособительных реакций целостного организма и составляющих его систем: пищеварительной, оборонительной, дыхательной, иммунной, кровеносной, половой, гормональной и т. д. Такой междисциплинарный характер исследований требует высокого уровня абстракции, отражающей наиболее общие закономерности роста, развития и жизнедеятельности организмов различной сложности, что в конечном счете должно было привести и привело к формированию нейрокибернетических принципов и методов структурно-функциональной организации живых систем. Поэтому неудивительно, что для раскрытия механизмов становления условных рефлексов еще в 1935 году в лаборатории лауреата Нобелевской премии И.П. Павлова был использован один из базовых принципов кибернетики и нейрокибернетики - принцип обратной связи (П.К. Анохин). Дальнейшее совершенствование аналитических методов исследований физических, биофизических, химических, биохимических, молекулярно-биологических и нейрофизиологических процессов жизнедеятельности привело к созданию теории функциональных систем [25], которая отличается от классической кибернетики в главном: обратная связь в живых системах используется не только и не столько для усиления и/или подавления уже существующих входных сигналов, а для опережающего формирования механизмов оценки качества результатов будущего действия, которые невозможно свести к одним лишь процедурам экстраполяции случайных процессов, положенным Н. Винером в основание кибернетики и хорошо изученным на данный момент.
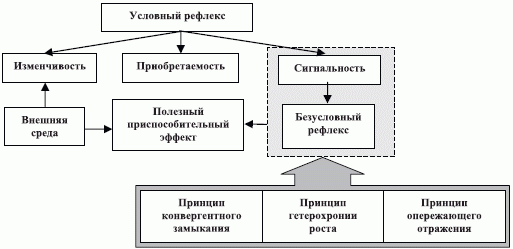
Базовым для теории функциональных систем является положение о том, что "единственным физиологическим принципом деятельности целого мозга, объясняющим поведение животных и человека, является принцип условного рефлекса [25]". Его справедливость вытекает из того факта, что в процессе обучения, без которого не обходится ни одна приспособительная деятельность организма, прежде всего должна быть установлена объективная, субъективная или даже волюнтаристская "причинно-следственная связь". Фактически должна быть построена своего рода "личная теория", формулирующая правила или алгоритм поведения индивидуума в конкретной обстановке. И только закрепив такую достаточно условную причинно-следственную связь между воздействием, именуемым "стимулом", и локальной, безусловно-рефлекторной, реакцией организма, можно рассматривать сформированный условный рефлекс в качестве "неделимой единицы физиологического проекта" и встраивать его в бол ее "сложный" целостный поведенческий акт, состоящий из последовательности локальных реакций, направленных на достижение той или иной цели поведения.
Здесь термин "условная связь" используется в расширенной трактовке, которая дополняет классическую Павловскую трактовку субъективным опытом отдельной особи, который может быть проигнорирован другими особями, особенно если их обучение протекает по принципу "делай как я". В частности, это может произойти, когда установленная отдельной особью "причинно-следственная связь" на самом деле является виртуальной и не имеет никакого отношения к реальным физико-химическим или молекулярно-биологическим законам взаимодействия с внешней средой. По И.П. Павлову термин "условный" используется в более узком смысле. Он подчеркивает, что данный тип реакции возможен только при воспроизводстве условий выработки причинно-следственной связи между одним из возможных стимулов и одной из возможных безусловных (врожденных, не приобретенных) физиологических реакций организма.
Приспособительные признаки условного рефлекса по И.П. Павлову (излагается по [25]):
- Приобретаемость или отсутствие врожденности этого типа реакций, что является главным критерием для выделения их в отдельную категорию приспособительных реакций организма.
- Изменчивость приобретенной нервной связи, которая постепенно теряется, как только становится неадекватной вновь сложившемуся комплексу внешних условий, в рамках которого утрачивается приспособительный эффект безусловного рефлекса.
- Сигнальность, которая предвосхищает дальнейший ход последовательно развивающихся событий внешнего мира.
Сигнальный характер условного рефлекса имеет глубокие молекулярно-биологические корни и является физиологическим проявлением общего для живых систем принципа опережающего отражения событий "неорганического" мира [25]. Этот принцип исходит из того, что даже простейшие биологические существа нашей планеты уже представляли собой системы (полу)открытого типа, которые связаны с окружающей средой через мембрану с избирательной проницаемостью рядом химических превращений, зарождающихся на мембране и продолжающихся целой серией многосвязных многостадийных ферментативных реакций внутри клеток и простейших организмов [60]. Исход такого взаимодействия живых систем с окру-
жающей средой априори никогда не известен, но негативный или даже трагический опыт такого взаимодействия можно учесть, а позитивный опыт воспроизвести только при условии повторяемости событий внешнего по отношению к организму мира. Поэтому для того чтобы организм смог вписаться в пространственно-временную структуру неорганического мира, у него должны быть механизмы опережающего отражения хотя бы повторяющихся в нем последовательных событий (рис. 4.5 [25]).
Из рис. 4.5 видно, если организм воспринимает некоторую последовательность событий как совокупность независимых воздействий, то он может реагировать на каждое из них только в отдельности и только постфактум. В этом случае у него нет возможности для активного противодействия внешним угрозам или для создания благоприятных условий эффективной "встречи" с биологически полезным событием "Д" во внешней среде (рис. 4.5-а). Напротив (рис. 4.5-б), если организм установил "причинно-следственную связь", гарантирующую ему более чем в 50 % случаев наступление события "Д" после того, как наступило событие "А", то у него есть время для подготовки условий встречи с биологически полезным событием "Д" или активного противодействия угрозе, проистекающей из такой встречи.
Таким образом, выработка условного рефлекса или, что одно и то же установление условной причинно-следственной связи между последовательными событиями внешней среды всегда сопровождается упреждающим синтезом молекулярно-биологических структур, адекватных условиям функционального взаимодействия с биологически важным событием. Но поскольку между структурой и функцией нет и не может быть взаимно однозначного соответствия, то встает вопрос о принципах и методах отбора адекватных молекулярно-биологических структур.
![Два типа реакций протоплазмы на последовательность внешних воздействий [25]](/EDI/14_06_16_2/1465856498-31733/tutorial/593/objects/4/files/04_05.gif)
Структурно-функциональное единство живых систем в теории функциональных систем представлено принципом гетерохронии роста и эмбрионального развития, который применим и к сложным техническим системам. Данный принцип исходит из того, что для выживания новорожденный должен обладать необходимым комплексом структурно-функциональных возможностей, который включает [25]:
- специфический рецепторный аппарат для восприятия экологических воздействий;
- средства доставки периферической информации в центральную нервную систему;
- управляемые (перестраиваемые) синаптические механизмы межней-рональных взаимодействий в центральной нервной системе, ответственной за интегрирование полноценного акта;
- исполнительный периферический аппарат с нервными окончаниями, обеспечивающий рабочий эффект системы;
- средства обратной афферентации, сигнализирующей о степени успешности каждого жизненно важного приспособительного действия новорожденного (афферентный - восходящий).
Экспериментальные и натурные данные говорят о том, что гете-рохронный и избирательный рост структур зародыша не направлен на равномерное развитие любого органа как целого. Он служит главной цели эволюции - наделению новорожденного полноценными и жизненно важными функциональными системами. В результате в развитии зародыша предпочтение отдается формированию жизненно важной функции, возможно, и в ущерб созданию того или иного органа как целого. Характерное проявление этого принципа - "шумы" сердца у новорожденных и в более позднем подростковом возрасте, которые не вызваны патологией сердечной мышцы.
В результате можно считать, что гетерохронность является базовым механизмом системогенеза, в том числе и сложных технических систем, в рамках которого предпочтение отдается избирательному и ускоренному развитию тех разнообразных по качеству и локализации структурных образований, которые, консолидируясь в целом, интегрируют полноценную функциональную систему, обеспечивающую новорожденному выживание [25].
Принцип опережающего отражения фактически ставит вопрос о механизмах трансформации системных макровзаимодействий на уровень молекулярных процессов и наоборот. Ответ на первую часть вопроса дает принцип конвергентного замыкания условного рефлекса, который был сформулирован П.К. Анохиным [25] в виде гипотезы, так как на тот момент отсутствовала достаточная нейрохимическая экспериментальная база. Тем не менее, этот принцип раскрыл схему перевода индифферентного
стимула в биологически значимый условный сигнал,что и является нейрофизиологическим итогом любого "обучения" организма. По П.К. Анохину, в логической схеме конвергентного замыкания на макроуровне задействованы два типа каналов поступления генерализованного возбуждения коры больших полушарий, по которым передается информация как об индифферентных, так и о безусловных раздражителях. Каналы первого типа проходят через лемнисковую систему и таламус к проекционным зонам коры, а второго типа - через ретикулярную формацию, связанную разветвленной сетью коллатералей с каналами первого типа [61]. Основная особенность возбуждений коллатерального ответвления в ретикулярную формацию состоит в том, что они являются опосредованными, множественными копиями возбуждений, направляемых в таламус и кору. Поэтому можно считать, что все подкорковые образования, хотя и с разной степенью пространственно-временной активности, постоянно задействованы в генерализованном возбуждении коры больших полушарий. Это является необходимым условием встречи на одном или группе корковых нейронов (конвергентных нейронов) требуемой комбинации пока еще индифферентного и биологически значимого возбуждений из (под)мно-жества возможных комбинаций. В их отборе (из множества всевозможных комбинаций), по П.К. Анохину, участвует сложный комплекс (нейро) физиологических механизмов функциональной системы.
Сама по себе встреча индифферентного и безусловного возбуждений на корковом нейроне также не является достаточным условием для трансформации такой первоначально разнесенной во времени и пространстве пары в устойчивую условно-рефлекторную связь, даже если эта пара часто воспроизводится в процессе жизнедеятельности организма. Согласно теории функциональной системы из всех таких повторяющихся пар отбираются только те, которые, будучи спроецированы на комплекс эфферентных (нисходящих) возбуждений, обеспечивают организму устойчивый, биологически полезный приспособительный эффект. При этом необязательно тождественное воспроизведение всего комплекса внешних и внутренних условий существования организма. Поэтому на последней стадии афферентного (восходящего) синтеза отбор и закрепление устойчивых комбинаций теперь уже условных и безусловных раздражителей осуществляется на уровне метаболических процессов конвергентных нейронов,где только и возможно взвешенное сопоставление оперативного опыта "(само)обучающейся" особи с опытом ее эволюционного и эмбрионального становления и развития.
Согласно [62] приобретенный особью опыт фиксируется на уровне молекулярных процессов клетки. Это нашло экспериментальное подтверждение в опытах с "каннибальными" планариями, которые обучались гораздо быстрее, получив с пищей информацию об оперативно накопленном опыте
через клеточный субстрат, "добытый" из сородичей с выработанным условным рефлексом. H. Hyden в своей теории молекулярной памяти [63] раскрыл роль РНК в процессе "обучения" эволюционно более поздних нервных клеток, одно только возбуждение которых приводит к специфическим структурным перестройкам нативного белка. По H. Hyden, схема "записи" информации в макромолекулярную память имеет следующий вид:
- в аксоплазме нервной клетки в зависимости от частоты возбуждения происходит перестройка РНК, приводящая к изменению состава или положения нуклеотидов в ее цепочке;
- изменение матричной РНК неизбежно приводит к изменению структуры синтезируемых на ее основе нативных белков, которые и являются хранителями полученной информации.
Схема "чтения" структурно представленной информации имеет тот же вид, но приходящие с той же частотой возбуждения теперь уже разрушают сформировавшийся комплекс (био)химических реакций. Именно этот эффект "перерегулирования" приводит к возбуждению нейрона, который стремится восстановить структуру нативных белков, регенерируя их по классической схеме: "транскрипция ДНК - матричная РНК - фермент - белок".
Современные исследования показали, что в межнейрональных и субнейрональных взаимодействиях решающую роль играют не столько пресинаптические трансформации пришедших возбуждений, сколько специфическая биохимическая активность веществ в составе постсинап-тической мембраны. Однако этот факт не изменяет в целом рассмотренную схему "чтения-записи" информации кодируемой структурой внутри-и внеклеточных белков.
Из приведенных данных видно, что в реальных нейронах:
- отбор комбинаций индифферентных и безусловных раздражителей, "подозреваемых" на существование между ними условной причинно-следственной связи, осуществляется многоуровневым, многостадийным процессом; этот процесс завершается в метаболизме конвергентных нейронов коры, где только и можно найти баланс между оперативным, генетическим и эволюционным опытом "(само)обучающейся" особи;
- необходимые условия отбора и закрепления устойчивых причинно-следственных связей в цепочках условных стимулов обеспечивает (на макроуровне) гиперпараллельная система пространственно-временной коммутации; ее адаптация (в темпе реального времени) осуществляется достаточно сложным комплексом механизмов функциональной системы;
- на ограниченных интервалах времени, диктуемых в основном доминирующей мотивацией, фиксация, закрепление и воспроизводство оперативного опыта особи осуществляется через динамические изменения субнейрональных молекулярных структур, которые синтезируются по консервативным законам эволюции и эмбрионального развития.
Таким образом, если организмы рассматривать как прототип систем, где успешно решены проблемы синтеза "ненадежными" методами и средствами из "ненадежных" компонент с "малым временем жизни", то в систематику синтеза нанометровой или супрамолекулярной вычислительной техники необходимо включить следующие базовые положения теории функциональных систем П.К. Анохина (рис. 4.6) [25]:
- Опережающее отражение действительности - это основная форма приспособления живой материи к пространственно-временной структуре неорганического мира, в котором само выживание обусловлено последовательным и повторяющимся характером событий, а активное отношение к событиям внешней среды - возможностью выбора из множества "стимулов" и множества допустимых реакций адекватных условных причинно-следственных связей.
- В живых системах итогом обучения является установление на молекулярно-биологическом уровне однозначного соответствия между единственным стимулом и единственно возможной в данном комплексе внешних условий реакцией, причем для фиксации такого однозначного соответствия достаточно использовать не аналитическую, а табличную форму его представления.
- При обработке информации в реальных нейронах и нейронных образованиях используется комплекс взаимно обусловленных структурно-параметрических методов ее хранения и преобразования, когда предыстория обработки закрепляется в некоторой молекулярно-биологической структуре, которая используется в дальнейших преобразованиях.
- Методами и средствами нейроподобной компиляции можно представить задание пользователя непосредственно в физико-химическом или супрамолекулярном операционном базисе.